Executive Neural Brief
Emotional Intelligence (EI) is frequently mischaracterized as a soft skill or personality trait. From a neurophysiological perspective, however, EI is a quantifiable measure of neural integration and efficiency. It represents the functional connectivity between the phylogenetically ancient limbic system—specifically the amygdala—and the evolutionarily recent prefrontal cortex (PFC). This hub examines the neuroanatomical substrate of emotional intelligence through three primary mechanisms: amygdala-PFC regulation, uncinate fasciculus connectivity, and the Real-Time Neuroplasticity™ framework for emotional regulation mastery.
Core Neurobiological Mechanisms
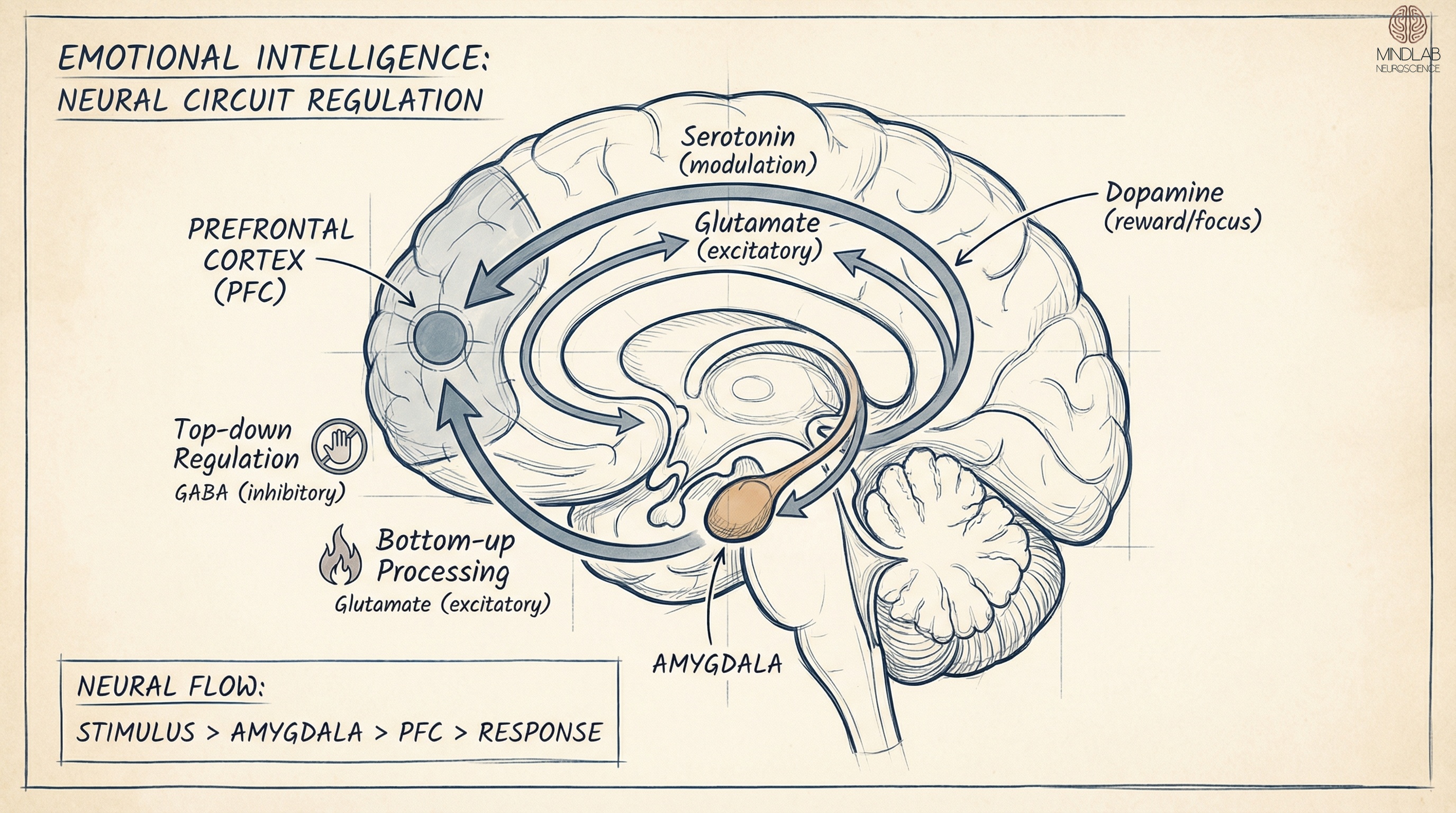
Amygdala-Prefrontal Cortex Regulation: The core of emotional processing resides in the amygdala, responsible for detecting salient stimuli, particularly threat-related inputs. In EI mastery, the Ventromedial Prefrontal Cortex (vmPFC) acts as the primary regulatory mechanism, exerting inhibitory “top-down” control over amygdala activation. High EI correlates with increased cortical thickness in the vmPFC and enhanced functional coupling between these regions, preventing “limbic hijack” and maintaining cognitive flexibility under stress.
Uncinate Fasciculus Connectivity: Structural connectivity between emotional centers (limbic system) and executive centers (frontal lobes) relies on the uncinate fasciculus white matter tract. Individuals exhibiting emotional intelligence mastery demonstrate higher myelination density within this tract, enhancing conduction velocity and reducing latency between emotional triggers and regulated behavioral responses. This structural efficiency is the neurobiological basis of “composure.”
Interoception and Somatic Awareness: Emotional self-awareness is mediated by the Anterior Insular Cortex (AIC), which integrates visceral afferent data with cognitive context. High EI individuals possess a highly active and differentiated AIC, allowing granular detection of emotional shifts before they escalate into overwhelming affect. This enables earlier initiation of regulation strategies within the emotional cycle.
Neuroscience-Based Implications
Emotional intelligence mastery represents not a static personality trait but a dynamic neurophysiological state achieved through deliberate reinforcement of top-down regulatory circuits. Modern humans operate with Paleolithic neural hardware in a digital environment, creating evolutionary mismatch where psychosocial “threats” trigger the same neurochemical cascade as physical dangers. Mastery requires cognitive override of obsolete survival protocols through metabolic optimization and HPA axis recalibration.
The Neuroanatomical Substrate of Emotional Intelligence
Emotional Intelligence (EI) is frequently mischaracterized as a soft skill or a personality trait. From a neurophysiological perspective, however, EI is a quantifiable measure of neural integration and efficiency. It represents the functional connectivity between the phylogenetically ancient limbic system—specifically the amygdala—and the evolutionarily recent prefrontal cortex (PFC). Mastery of emotional intelligence is not merely psychological; it is the result of optimized cytoarchitecture and robust white matter integrity within specific neural circuits.
The core of emotional processing resides in the amygdala, a bilateral structure within the temporal lobes responsible for detecting salient stimuli, particularly threat-related inputs. In an untrained or dysregulated brain, the amygdala operates via a “bottom-up” processing mechanism. It receives sensory input from the thalamus and initiates the hypothalamic-pituitary-adrenal (HPA) axis response before the conscious mind can intervene. This results in the release of cortisol and norepinephrine, triggering sympathetic nervous system arousal—the fight-or-flight response.
In the context of EI mastery, the Ventromedial Prefrontal Cortex (vmPFC) acts as the primary regulatory mechanism. The vmPFC exerts inhibitory control over the amygdala. This “top-down” regulation allows for the modulation of affective states. High EI correlates with increased cortical thickness in the vmPFC and enhanced functional coupling between the vmPFC and the amygdala. Mastery involves the rapid recruitment of the vmPFC to dampen amygdalar activation, thereby preventing “limbic hijack” and maintaining cognitive flexibility under stress.

The Uncinate Fasciculus and Signal Transduction
Structural connectivity is as critical as functional activation. The communication between the emotional centers (limbic system) and the executive centers (frontal lobes) relies heavily on the uncinate fasciculus. This hook-shaped white matter tract connects the amygdala and hippocampus with the orbitofrontal cortex (OFC). The integrity of this tract, measured via Diffusion Tensor Imaging (DTI) as fractional anisotropy, is a biological predictor of emotional regulation capabilities.
Individuals exhibiting emotional intelligence mastery demonstrate higher myelination density within the uncinate fasciculus. Increased myelination enhances the conduction velocity of neural action potentials. This allows the prefrontal cortex to receive emotional data faster and transmit inhibitory signals back to the limbic system with greater efficiency. Consequently, the latency period between an emotional trigger and a regulated behavioral response is significantly reduced. This structural efficiency is the neurobiological basis of “composure.”
Interoception: The Role of the Anterior Insular Cortex
A critical, often overlooked component of emotional intelligence is self-awareness, or the ability to perceive one’s own physiological state. This process, known as interoception, is mediated by the Anterior Insular Cortex (AIC). The insula receives visceral afferent data—heart rate, gastric tension, temperature—and integrates this somatic information with cognitive context.
In the neurophysiology of EI mastery, the AIC serves as a hub for the “somatic marker hypothesis.” It translates bodily sensations into emotional awareness. High-resolution fMRI studies suggest that individuals with high EI possess a highly active and differentiated AIC. This allows for the granular detection of emotional shifts before they escalate into overwhelming affect. By consciously recognizing the physiological precursors of emotion (e.g., slight tachycardia or muscle tension), the executive brain can initiate regulation strategies earlier in the emotional cycle.
Neurochemical Modulation: Serotonin and Dopamine Pathways
The synaptic transmission of emotional data is governed by specific neurotransmitter systems. Serotonin (5-HT) plays a pivotal role in the “braking” mechanism of emotional responses. Adequate serotonergic transmission in the raphe nuclei and its projections to the PFC is essential for impulse control and the inhibition of aggressive or reactive behaviors. Deficits in serotonergic signaling are linked to a decoupling of the amygdala-PFC circuit, leading to emotional volatility.
Conversely, the mesocorticolimbic dopamine system influences motivation and reward prediction. Dopamine is not merely a pleasure molecule; it is a learning signal. In the context of EI, dopamine facilitates cognitive flexibility—the ability to shift perspective and update emotional responses based on new information. Mastery involves a balanced dopaminergic tone that supports goal-directed behavior without succumbing to immediate gratification or impulsive reactivity.
Furthermore, the balance of norepinephrine is critical. While essential for alertness, excessive noradrenergic activity in the locus coeruleus impairs prefrontal cortical function. This phenomenon, known as the “inverted-U” curve of arousal, dictates that optimal emotional intelligence operates within a specific window of catecholamine release. Mastery entails the physiological capacity to maintain norepinephrine levels within this optimal window, preserving working memory and executive function even during high-pressure scenarios.
Synaptic Plasticity and the Mechanism of Mastery
The development of emotional intelligence is underpinned by neuroplasticity—specifically, Long-Term Potentiation (LTP). LTP is the persistent strengthening of synapses based on recent patterns of activity. When an individual engages in cognitive reappraisal (reframing a negative event), they activate the lateral Prefrontal Cortex (lPFC). Repeated activation of the lPFC in conjunction with the downregulation of the amygdala strengthens the synaptic connections between these regions.
This is the mechanism of Hebbian learning: “neurons that fire together, wire together.” Through consistent practice of emotional regulation techniques, the brain physically alters its circuitry. Dendritic arborization increases in the prefrontal regions, and synaptic efficacy improves in the inhibitory pathways leading to the limbic system. Therefore, emotional intelligence mastery is not a static trait but a dynamic neurophysiological state achieved through the deliberate, repetitive reinforcement of top-down regulatory circuits.
The Phylogenetic Mismatch: Paleolithic Hardware in a Digital Environment
To understand the neuroscientific basis of emotional intelligence (EI), one must first interrogate the evolutionary architecture of the human brain. We operate with a neural apparatus designed for the late Pleistocene era, yet we function within a high-latency, digital sociopolitical environment. This discrepancy is known as the evolutionary mismatch theory.
The human limbic system, specifically the amygdala, evolved as a rapid-response threat detection mechanism. In ancestral environments, threats were physical and immediate—predation, inter-tribal violence, or environmental hazards. The physiological response (activation of the HPA axis: Hypothalamic-Pituitary-Adrenal) was binary and acute. Cortisol and adrenaline flooded the system to facilitate fight or flight, followed by a period of metabolic recovery.
In the modern context, “threats” are rarely physical but predominantly psychosocial—status anxiety, micro-aggressions, or information overload. The amygdala, lacking the sophistication to distinguish between a saber-toothed tiger and a hostile email, initiates the same neurochemical cascade. Consequently, modern humans exist in a state of chronic, low-grade HPA axis activation. Emotional intelligence mastery, therefore, is not merely a soft skill; it is the cognitive override of an obsolete survival protocol.
The Metabolic Cost of Top-Down Processing
Emotional regulation is metabolically expensive. The brain, while comprising only 2% of body mass, consumes approximately 20% of the body’s glucose and oxygen. This energy distribution is not uniform. The prefrontal cortex (PFC)—the seat of executive function, impulse control, and emotional regulation—is phylogenetically the newest and metabolically the most demanding region.
Conversely, the limbic system (bottom-up processing) is energy-efficient and rapid. Reacting with anger or fear requires minimal metabolic overhead because these neural pathways are highly myelinated and evolutionarily entrenched. Exercising emotional intelligence requires “top-down” processing: the PFC must inhibit the limbic impulse, reframe the context, and generate a nuanced response.
This process, known as cognitive inhibition, depletes glucose reserves. This biological reality explains the phenomenon of “ego depletion,” where self-control acts as a finite resource. When an individual is fatigued, hypoglycemic, or cognitively overloaded, the metabolic cost of EI becomes prohibitive, and the brain defaults to energy-saving, limbic-driven behaviors. Mastery requires optimizing metabolic substrates to support the heavy lifting of the PFC.
The Agricultural Inflection: Chronic Stress and Cortisol Dysregulation
The Neolithic Revolution—the transition from hunter-gatherer societies to agricultural civilizations—fundamentally altered the human stress profile. In the Paleolithic, stressors were acute, episodic, and physically resolved. The encounter with a predator or rival group triggered a massive HPA axis response, flooding the system with cortisol and catecholamines. This was followed by either death or survival, with survivors experiencing a rapid return to baseline cortisol levels.
Agriculture introduced a novel stressor: chronic, unresolvable psychological stress. The farmer faced existential threats that could not be fought or fled from—crop failure, debt, hierarchical oppression. These threats activated the HPA axis continuously but without the metabolic resolution that physical action provides. Over generations, this chronic activation rewired the stress response system.
Modern emotional intelligence operates within this inherited framework. The contemporary brain is predisposed to cortisol dysregulation—either hypercortisolemia (chronic elevation) or a blunted cortisol awakening response (adrenal fatigue). Both phenotypes impair the PFC’s ability to exert top-down control over the amygdala. High cortisol degrades hippocampal tissue, impairing memory consolidation and contextual processing. Low cortisol results in insufficient arousal, manifesting as apathy and emotional numbness.
Emotional intelligence mastery, therefore, requires the active recalibration of the HPA axis. This is not achieved through willpower alone but through targeted interventions: circadian alignment, metabolic optimization, and the deliberate cultivation of acute, resolvable stressors (e.g., cold exposure, high-intensity interval training) that restore the brain’s capacity for rapid stress onset and offset.
The Dopaminergic Deficit: Reward Insensitivity and Anhedonia
The shift to agricultural societies also disrupted the dopaminergic reward system. In the hunter-gatherer context, effort and reward were tightly coupled in time. The successful hunt delivered an immediate dopamine spike, reinforcing the behavior within hours. Agriculture, by contrast, introduced a “delayed return” environment: plant seeds in spring, harvest in autumn. Effort and reward were decoupled by months.
This created a selection pressure for individuals capable of tolerating dopaminergic ambiguity—those who could sustain motivation despite extended periods without reward. However, this adaptation came at a cost. Modern populations descended from agricultural lineages exhibit reduced D2 receptor density in the striatum, a marker of reward insensitivity.
In the context of emotional intelligence, this manifests as anhedonia—the inability to experience pleasure from social connection or self-regulation. Individuals with low D2 receptor density struggle to derive intrinsic motivation from the practice of emotional regulation. The dopamine “hit” from successfully managing a difficult emotion is insufficient to reinforce the behavior, leading to relapse into reactive patterns.
Mastery requires the restoration of dopaminergic sensitivity. This is achieved not by increasing dopamine levels (which leads to tolerance and downregulation) but by recalibrating the reward system through intermittent reinforcement, novelty exposure, and the strategic use of immediate feedback loops that restore the brain’s sensitivity to small, incremental wins.
Mechanisms of Real-Time Neuroplasticity™
The foundational premise of this protocol is that the brain is not a static storage device, but a dynamic architecture that remodels itself instantly in response to cognitive demand. While traditional training focuses on muscular output, Real-Time Neuroplasticity™ (RTN) targets the neural input and processing stages.
To achieve high-performance adaptation, we leverage three distinct biological mechanisms: Directed Neuroplasticity, Synaptic Pruning, and Myelination. Understanding these mechanisms transforms training from simple repetition into structural neural engineering.
Directed Neuroplasticity: The Architecture of Intent
Neuroplasticity is the brain’s ability to reorganize itself by forming new neural connections. In a passive state, the brain adapts to whatever environment it is placed in. In a high-performance context, we utilize Directed Neuroplasticity—the deliberate forcing of neural adaptation through specific, high-load cognitive demands.
This mechanism relies on the Hebbian principle: “Cells that fire together, wire together.”
When a specific cognitive task is paired with a physical action, the neurons involved in that chain facilitate chemical changes at the synapse (Long-Term Potentiation). Repeated, focused firing strengthens these connections, moving a skill from a temporary working memory state to a physical neural structure.
The RTN Distinction: Random repetition creates “noise” in the neural map. Directed Neuroplasticity requires high-fidelity signal input. If the cognitive intent is vague, the wiring is weak. If the cognitive intent is precise, the structural change is robust.
Synaptic Pruning: Signal Optimization
If Directed Neuroplasticity is the architect building new roads, Synaptic Pruning is the demolition crew removing the old ones to prevent traffic jams.
The brain operates on an economy of energy. Maintaining neural pathways that are inefficient, contradictory, or rarely used is metabolically expensive. Through a process involving microglia (the brain’s immune cells), the brain eliminates weak synaptic connections. This process is often summarized as: “Use it or lose it.”
In the context of RTN, pruning is essential for eliminating “neural noise”—the micro-hesitations, incorrect movement patterns, and delayed processing speeds that hinder elite performance.
The Efficiency Loop: By strictly enforcing correct neural pathways and refusing to energize incorrect ones, we accelerate the pruning of bad habits. As the brain eliminates competing neural options, decision-making speed increases because the signal no longer has to navigate a clutter of alternative choices.
Myelination: The Velocity Factor
Once a pathway is built (Directed Neuroplasticity) and cleared of noise (Synaptic Pruning), it must be insulated for speed. This is the function of Myelination.
Myelin is the fatty sheath that wraps around neural axons, produced by glial cells called oligodendrocytes. Its primary function is to increase the conduction velocity of electrical signals (action potentials). An unmyelinated axon transmits signals at approximately 1-2 meters per second. A fully myelinated axon can reach speeds of up to 120 meters per second—a 100x increase.
In the RTN framework, myelination is not a passive process that occurs over years. It can be accelerated through high-frequency, high-intensity neural firing. When a neural circuit is activated repeatedly and with high precision, it triggers oligodendrocyte precursor cells to differentiate and wrap the axons in myelin.
The Performance Payoff: Myelination transforms a conscious, effortful skill into an unconscious, high-speed reflex. This is the biological basis of “muscle memory” or “automaticity.” In emotional intelligence, this means that regulated responses become instantaneous rather than requiring deliberate cognitive effort.
Frequently Asked Questions
What is the fundamental neuroanatomical difference between high and low emotional intelligence?
The primary neuroanatomical distinction lies in the structural and functional connectivity between the amygdala (emotional processing center) and the prefrontal cortex (executive control center). Individuals with high emotional intelligence demonstrate: (1) Increased cortical thickness in the ventromedial prefrontal cortex (vmPFC), (2) Enhanced functional coupling between vmPFC and amygdala during emotional tasks, (3) Higher myelination density in the uncinate fasciculus white matter tract connecting these regions, and (4) Greater activation and differentiation in the anterior insular cortex for interoceptive awareness. These structural advantages enable faster, more efficient top-down regulation of emotional responses.
How does chronic stress impair emotional intelligence at the neurobiological level?
Chronic stress induces structural and functional changes that directly compromise emotional intelligence. Elevated cortisol levels: (1) Reduce dendritic branching in the prefrontal cortex, impairing executive function and top-down regulation, (2) Increase dendritic arborization in the amygdala, enhancing emotional reactivity, (3) Degrade hippocampal tissue, impairing contextual memory and the ability to differentiate between similar emotional situations, and (4) Reduce myelination in the uncinate fasciculus, slowing communication between emotional and regulatory centers. These changes create a neurobiological state where emotional reactions become faster and stronger while regulatory capacity becomes slower and weaker, fundamentally altering the brain’s emotional architecture.
Can emotional intelligence be improved through specific neural exercises?
Yes, emotional intelligence can be enhanced through targeted neural exercises that leverage neuroplasticity mechanisms. Effective interventions include: (1) Cognitive reappraisal training to strengthen prefrontal-amygdala connectivity, (2) Interoceptive awareness exercises to enhance anterior insular cortex differentiation, (3) Mindfulness practices to increase gray matter density in prefrontal regions, (4) Social cognition training to improve mirror neuron system function, and (5) Stress inoculation protocols to recalibrate HPA axis responsiveness. These exercises work by inducing Long-Term Potentiation in regulatory pathways, promoting myelination of the uncinate fasciculus, and optimizing neurotransmitter balance for emotional regulation.
What role do neurotransmitters play in emotional intelligence?
Neurotransmitters serve as the chemical messengers that modulate emotional intelligence circuits: (1) Serotonin enhances impulse control and emotional stability through projections from raphe nuclei to prefrontal cortex, (2) Dopamine facilitates cognitive flexibility and reward-based learning in emotional contexts, (3) Norepinephrine maintains optimal arousal levels for emotional regulation (following an inverted-U curve), (4) GABA provides inhibitory control over amygdala hyperactivity, and (5) Oxytocin enhances social bonding and trust perception. Emotional intelligence mastery requires balanced neurotransmitter systems where each chemical operates within optimal ranges rather than maximal levels, creating a neurochemical environment conducive to regulated emotional responses.
How does the “somatic marker hypothesis” explain emotional decision-making?
The somatic marker hypothesis proposes that emotional signals from the body (somatic markers) guide decision-making by creating physiological “gut feelings” about potential outcomes. This process involves: (1) The anterior insular cortex detecting bodily changes associated with emotional states, (2) The ventromedial prefrontal cortex associating these somatic signals with specific scenarios, (3) The amygdala tagging emotionally significant memories with somatic markers, and (4) The body generating physiological responses (heart rate changes, muscle tension) when similar scenarios are encountered. In emotional intelligence mastery, individuals develop heightened sensitivity to these somatic markers, allowing them to access emotional wisdom before conscious cognitive processing completes, leading to more adaptive decisions in complex social situations.
Selected Scientific References
- Damasio, A. R. (1994). Descartes’ Error: Emotion, Reason, and the Human Brain. Putnam Publishing.
- Davidson, R. J., & Irwin, W. (1999). The functional neuroanatomy of emotion and affective style. Trends in Cognitive Sciences, 3(1), 11–21.
- Lieberman, M. D. (2007). Social cognitive neuroscience: A review of core processes. Annual Review of Psychology, 58, 259–289.
- Ochsner, K. N., & Gross, J. J. (2005). The cognitive control of emotion. Trends in Cognitive Sciences, 9(5), 242–249.
- Pessoa, L. (2008). On the relationship between emotion and cognition. Nature Reviews Neuroscience, 9(2), 148–158.
About Dr. Sydney Ceruto
Dr. Sydney Ceruto is a Neuroscientist and Brain Performance Strategist specializing in neurological re-engineering for elite individuals navigating high-stakes environments. As the founder of MindLAB Neuroscience and the pioneer of Real-Time Neuroplasticity™, she translates clinical neurobiology into decisive competitive advantages for tech innovators, professional athletes, entertainers, and private families worldwide. Dr. Ceruto holds dual PhDs in Behavioral & Cognitive Neuroscience from New York University and dual Master’s degrees in Clinical Psychology and Business Psychology from Yale University. She is the author of The Dopamine Code, published by Simon & Schuster.